1.-
Introducción
2.-
Adaptaciones a la inmersión.
3.-
Vida a grandes profundidades.
5.-
Medicina hiperbárica
4.-Efecto
de la hiperbaria sobre el organismo humano
1.-
Introducción
Entendemos por hiperbarismo, cualquier alteración que se
produce como consecuencia de la exposición a una presión ambiental aumentada,
habitualmente por la exposición brusca a una presión aumentada o por un
incremento significativo de la presión.
Las variaciones de presión son mucho
más importantes en el medio acuático que en la montaña. Es necesario ascender
por encima de los 5.800 m para disminuir a la mitad la presión atmosférica, por
contra,
basta descender a 10 m de profundidad en el agua para duplicar la
presión ambiental. El aumento de presión modifica el comportamiento de algunos
sistemas orgánicos, en especial en lo que concierne a los gases contenidos en
el organismo, en los que produce variaciones físicas, o de volumen, y
variaciones químicas. Para entender estas variaciones y las modificaciones orgánicas
que provocarán, debemos tener presente las siguientes leyes físicas:
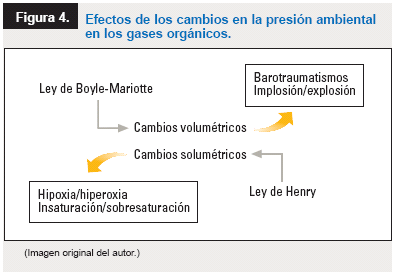
- La
ley de Boyle-Mariotte
establece que el producto del volumen que ocupa un gas y la presión a que
está sometido se mantiene siempre constante. Por consiguiente, si se
aumenta o disminuye la presión, disminuirá o aumentará en proporción inversa
el volumen.
P*V=
k, cuando T permanece constante.
Luego:
P1*V1= P2*V2
Donde:
P son las
presiones inicial (1) y final (2)
V son los
volúmenes inicial (1) y final (2)
§
Según la ley
de Henry, todos los gases tienden a disolverse en los líquidos en
contacto, de un modo proporcional a la presión parcial del gas a nivel de la
superficie del líquido. En otras palabras, la solubilidad de un gas en un
disolvente es proporcional a la presión del gas sobre el líquido.
S
= ks * P
Donde:
P
es la presión parcial del gas.
S
es la concentración del gas (solubilidad).
ks
es la constante de Henry, que depende de la naturaleza del gas, la temperatura
y el líquido.
2.- Adaptaciones a la inmersión.
E10 metros de profundidad.
Esto supone, como es natural, la existencia de enormes presiones, que llegan a
sobrepasar las 1000 atmósferas en las mayores profundidades oceánicas, que son
de 11000 metros.
n los océanos, la presión que
ejerce la atmósfera aumenta en razón de una atmósfera por cada
La profundidad media de los océanos es de 4000 metros y está
sometida, por lo tanto, a una presión de 400 atmósferas. La presión hasta la
que se han encontrado la mayor cantidad de organismos marinos es de 600
atmósferas, ya que en los fondos mayores la cantidad disminuye y se localizan
principalmente algunos invertebrados y bacterias.
A modo de noción básica la distribución animal que encontramos en
los océanos es la siguiente. Los animales de aguas intermedias a profundidades
de entre 200 a 800 metros migran a aguas menos profundas cada noche para
alimentarse. Estas son conocidas como “migraciones verticales” y ocurren en
todo el mundo a gran escala. Los animales de estas profundidades se conocen
como “mesopelágicos”. Los animales que viven a profundidades mayores a 1000 m
se conocen como “batipelágicos”, lo que quiere decir “profundidad” y “vida
libre”. Están entre dos aguas, típicamente a una larga distancia del fondo.
Otros grupos de animales que también están entre dos aguas, pero mucho más
cerca del fondo, alimentándose de animales asociados al fondo marino, son los
conocidos como “bentopelágicos” (el término “demersal” se usa también para
describir a estos animales). Los animales que viven sobre (o en) el fondo se
conocen como “bénticos”, lo que significa “que viven en el fondo”. Aquellos que
viven en la superficie forman la “epifauna” y aquellos que están enterrados en
la arena o el fango se conocen como “infauna”.

Gráfica 1: Capas oceánicas.
El paso por los distintos niveles de distribución va a exigir una
serie de adaptaciones enfocadas a la inmersión, en este caso destacaremos
aquellas que guardan relación con los cambios barométricos, distinguiendo las
distintas necesidades de los organismos, puesto que estas serán diferentes en
función de que se requiera migraciones superficiales, vida permanente en el
fondo, etc.

Respuesta respiratoria.
La aeremia ocurre cuando el aire que la sangre ha absorbido de los pulmones,
mientras el animal estaba sumergido bajo alta presión, se va soltando como
burbujas durante el ascenso cuando la presión vuelve a bajar. Ballenas y focas
evitan esto al dejar que los pulmones se compriman durante la primera parte de
la sumersión. Por lo tanto no queda aire que pueda ser absorbido en la sangre.
La capacidad de renovación
de las inspiraciones es mucho mayor, llegando a estar entre el 80% y el 90% del aire de los pulmones frente al 10% o
20% de la mayoría de los mamíferos terrestres.

Así
mismo es de destacar la tendencia a presentar cierta resistencia a acumular
dióxido de carbono en los tejidos, pues es la acumulación de éste, en lugar de
la falta de oxígeno, lo que desencadena la respuesta respiratoria involuntaria.
También encontramos la
disminución del reflejo de Hering-Breuer (este consiste en que el estiramiento
de receptores a nivel pulmonar ante una inspiración profunda lleva
secundariamente a tratar de exhalar). Pero como al descender el volumen
pulmonar se reduce por efecto de la ley de Boyle-Mariotte, se reduce el
estiramiento y por ende elimina la necesidad de exhalar, por el contrario al aumentar el volumen retorna el deseo de
respirar.
Otro mecanismo consiste en producir el colapso torácico y pulmonar a
gran profundidad (el aire pasa de los pulmones a la tráquea, lo que reduce la
absorción de nitrógeno, que es tóxico), así como la transmisión rápida del nitrógeno de la sangre a los pulmones al
final de la inmersión. Todo ello combinado con un metabolismo cercano al
anaeróbico, por glucólisis, produciendo ácido láctico.

La cantidad de oxígeno necesaria para una sumersión larga no se
acumula en los pulmones sino en la gran cantidad de mioglobina en los músculos
de los animales, que es capaz de almacenar hasta
el 50% del oxígeno. La proporción de glóbulos rojos
es superior y la mioglobina está más concentrada, dando lugar a un color muy
oscuro en los músculos.
Recordemos que en
todos los mamíferos, la mioglobina retiene el oxigeno en la célula en reposo y
lo suelta sólo en una actividad muscular intensa, cuando la hemoglobina liberó
su oxigeno.
El sulfuro de hidrógeno, abundante en el entorno oceánico, es un
compuesto muy tóxico que inhibe la respiración al bloquear los lugares donde la
molécula de hemoglobina se une con el oxígeno, Riftia por ejemplo, dispone de una gran
cantidad de sangre (más del 30% del volumen de su cuerpo), y su hemoglobina
además de poseer una alta afinidad por el oxígeno tiene un sitio específico de
fijación para el sulfuro, pudiendo transportar ambas moléculas simultáneamente.
Respuesta cardiovascular.
Un proceso común en mamíferos marinos,
es la redistribución del flujo sanguíneo preferentemente a los órganos vitales, como los pulmones y el
corazón, en detrimento del aparato digestivo, y reducción de
la frecuencia cardiaca (bradicardia).
Esta respuesta se considera que está
dirigida a conservar O2. Inmediatamente después de sumergirse en el
agua se produce la bradicardia de inmersión o reflejo de zambullida, de origen
vagal, pues desaparece administrando atropina. Es independiente de la
bradicardia que se produce por la apnea simple. Se piensa que es debida al
contacto del agua con las mucosas de las fosas nasales y de la boca, siendo un
reflejo ancestral de nuestro supuesto pasado acuático. Puede provocarse
simplemente introduciendo la cara del sujeto en un recipiente con agua, aunque
es mayor si el sujeto se encuentra en apnea.
Durante la inmersión también se
produce una vasoconstricción periférica, fenómeno muy pronunciado en los
mamíferos acuáticos, en los que llegan a desaparecer los pulsos periféricos. En
respuesta a la apnea, en el ser humano se produce una contracción del bazo,
transfiriendo a la circulación hasta el 50% de la sangre que contiene, lo cual puede
permitir prolongar la duración de la apnea.
La respuesta vagal produce una
bradicardia sinusal, disminución de amplitud de la onda P, la onda T se hace
muy picuda y el QRS no se modifica. Pueden aparecer cortos periodos de bloqueo
A-V, escapes nodales y extrasístoles.
Cuando se emerge al final de la apnea
aumentan los niveles sanguíneos de ácido láctico, similares hasta ese momento a
los del sujeto en situación de reposo. Este fenómeno se considera debido a la
liberación de ácido láctico por el músculo durante la hipoxia, que pasa de
forma brusca al torrente circulatorio cuando desaparece la vasoconstricción
periférica.
Los mamíferos marinos,
tienen un volumen de sangre mayor al de los mamíferos de tamaño y peso similar,
y una capacidad mayor para almacenar oxígeno en la sangre y en los tejidos
musculares.

Vejiga natatoria.
La vejiga natatoria es un
órgano de flotación exclusivo de los peces y tiene su origen en un divertículo
del esófago. En algunas especies, se mantiene todavía la conexión de la vejiga
con el esófago a través del conducto neumático. Dichas vejigas reciben el
nombre de fisóstomas
y las presentan los peces “primitivos”.
Si seguimos la evolución, podemos constatar como progresivamente este
conducto de comunicación se ha ido reduciendo hasta llegar a la total
independencia del digestivo con las vejigas (fisoclistas) que son completamente
cerradas (estilo globo), y que las presentan los peces “modernos” tales como la
popular merluza o el bacalao.
Debemos decir que no todos
los peces presentan vejiga natatoria y que muchos de ellos, no ha sido por una
pérdida secundaria debido a su modo de vida, sino que nunca su grupo la
presentó durante la evolución como es el caso de los peces cartilaginosos.
Éstos solucionan el problema de la flotabilidad incorporando depósitos grasos
en el hígado, concretamente escualeno, que puede llegar a representar el 30%
del peso del animal.
El tamaño de la vejiga
natatoria es variable en los peces. La norma general nos dice que en las
especies marinas puede llegar a representar el 5% del peso del animal, mientras
que en los de aguas continentales puede llegar hasta el 7%, siempre en función
de la densidad.
Después de haber visto la
anatomía descriptiva a vista de pájaro de este órgano de flotación, es momento
de adentrarnos en su funcionamiento, es decir, en los procesos fisiológicos que
lleva asociados.
El mantenimiento de un
determinado volumen de gas en el interior de la vejiga natatoria con sus
paredes colapsables supone dos problemas básicos:
§
Segregar gas activamente en
contra de un gradiente de presión parcial muy elevado.
§
Controlar rápida y
eficazmente la presión interna de los gases para ajustar los cambios de
densidad del animal asociados a una variación de la profundidad.
La secreción de gas se
produce a nivel de una estructura especializada llamada glándula de gas
que está formada por un epitelio secretor dispuesto en serie a un conjunto de
capilares arterio-venosos (rete-mirabile) que actúan como un sistema de
intercambiador en contra corriente.

Los mecanismos que
intervienen en la cesión de los gases a la vejiga son:
1.
Pérdida de afinidad de la
hemoglobina por el oxígeno, causados por una masiva producción de ácido láctico
en el epitelio de la glándula de gas.
2.
Efecto salado. La enorme
secreción de lactato en el epitelio disminuye la solubilidad de los gases y
causa su desplazamiento de la fase líquida a la fase gaseosa.
3.
Retención en la
rete-mirabile. Se establece un intercambio en contracorriente de solutos y
gases, actuando como trampa de lactato y de otros solutos, con un efecto
multiplicador, provocando un aumento de la presión parcial de oxígeno y de la
de nitrógeno hasta decenas de atmósferas.
No sirve de nada introducir
gas dentro de la vejiga si ésta no es capaz de retenerlo, por ello existen dos
mecanismos:
1.
Gracias al efecto
multiplicador en contra corriente se evita la pérdida de gas hacia el torrente
circulatorio.
2.
Impermeabilización de la pared
de la vejiga:
§
cara interna está
recubierta de una secreción rica en fosfolípidos que favorecen la
solubilización y retención del gas.
§
cara externa está
recubierta de un depósito de cristales de guanina que contribuyen a disminuir
la difusión en la pared de la vejiga.
La eliminación de gas de la
vejiga se produce a través de un espacio en la pared de la misma que recibe el
nombre de ventana oval
y que se halla en comunicación con el sistema circulatorio. Normalmente, los
capilares quedan separados del espacio en el que se encuentra el gas por un
esfínter muscular, que al relajarse permite que el gas a gran presión de la
vejiga pase por simple difusión a la sangre donde se encuentra a presiones
similares a la atmosférica. En el caso de los fisóstomos, tenemos la
posibilidad de su eliminación a través del esófago, ya que recordemos
presentaban un conducto neumático que lo comunicaba con la vejiga con lo cual
se simplifica mucho el mecanismo.
El sistema nervioso autónomo
es el responsable del control de la presión interna de la vejiga natatoria a
través de la acción de sus divisiones: simpática (eliminación) y parasimpática
(secreción). Estas acciones se desencadenan a través de señales propioceptoras
(internas) que informan de la tensión en la pared de la propia vejiga.
Cuando los peces se alejan de la superficie colocan gas en su vejiga, ya que
al aumentar la presión debido a la profundidad, la vejiga disminuiría su tamaño
y el pez aumentaría su peso específico por lo que iría hundiéndose cada vez más
hasta llegar al fondo. Cuando un pez está en zonas profundas y quiere acercarse a la superficie
no tiene más remedio que reducir el gas de su vejiga, ya que al ascender disminuye
su presión hidrostática sobre ella con lo que acabaría por dilatarse
enormemente y estallaría. Esto puede observarse cuando se pesca un pez a
profundidad y se lleva rápidamente a la superficie. Cuando los pescadores
quieren mantener vivo uno de estos peces de profundidad, inmediatamente después
de sacarlo le pican un costado con una aguja hipodérmica gruesa, para facilitar
la salida de los gases de la vejiga, y es común que los peces así tratados
puedan seguir viviendo para cultivarlos
Con respecto a los
organismos que viven en los fondos marinos a grandes profundidades, los
mecanismos están orientados a soportar la presión más que a prepararse ante
cambios de esta, dado que no suelen ascender a la superficie. Es por ello que
las estrategias son permanentes, y se manifiestan a niveles morfológico,
etológico o metabólico, tanto para dar repuesta a los efectos directos como
indirectos de las altas presiones.
La consecuencia más
directa de la presión, insta a los organismos a igualar la presión interna con
la del medio, el hecho de que se trabaje siempre a la misma presión, descarta
mecanismos cambiantes de presión, como puede ser la vejiga natatoria, carente
en peces abisales, por lo que a esta profundidad se recurre a equiparar el
medio interno al externo, reduciendo la masa ósea y muscular de los organismos,
al tiempo que se aumenta la proporción de líquidos del cuerpo, dado que estos
son prácticamente incomprensibles. La mayoría de los peces
abisales son pequeños y poseen cuerpos blandos y huesos pequeños, debido en
parte a la ausencia de calcio, necesario para la formación de espinas, y de
vitamina D, necesaria para unos huesos consistentes. Otro ejemplo en este
sentido, es la carencia de estructuras internas duras (en pulpos y calamares). Una
forma de equiparar cuerpo y entorno, es por medio del incremento de los niveles de ácidos grasos insaturados.
Con respecto a la
pérdida de masa muscular, lo que a priori puede parecer una desventaja, las
condiciones del entorno, como la tranquilidad y el reposo de
las aguas de los profundos abismos del mar, hacen que los seres abisales no
tengan que contrarrestar la acción de las corrientes y, mucho menos, la del
oleaje, que no se dejan sentir. Así mismo, a medida que aumenta la presión,
aumenta la densidad y se gana empuje, hecho que se explica fácilmente si
comparamos la superficie de las alas de las aves, con las aletas de los peces,
y en última instancia las aletas de peces superficiales con peces abisales,
encontrándonos que a medida que descendemos la proporción de los apéndices se
reduce, así mismo
el uso de aletas flexibles disminuye el gasto energético, no
así las largas y rígidas que aumentan el costo en un 25% aproximadamente.
Otra forma de adaptarse a las grandes presiones es modificar la
morfología, dando lugar a formas aplanadas, dorsolventralmente deprimidas o
serpentiformes. Ejemplos en este sentido, son las rayas y lenguados, o las
morenas como representantes de las formas serpentiformes. De igual modo, los
organismos bentónicos, tienen la necesidad de no hundirse en el sedimento, por
lo que pueden tener apéndices corporales (Equinodermos); espículas en forma de
raíz (Hexactinélidas), o un pedúnculo más o menos largo (Octocorallarios,
Ascidias). Este aspecto es menos problemático para especies pequeñas
(Foraminíferos, pequeños Moluscos).
Además nos encontramos en un medio donde los productores se restringen a las bacterias, y
en el que las fuentes de alimento proceden bien de la depredación, o bien de
restos de la superficie, cuya caída es más rápida cuanto más aumenta la
presión, ya que ésta contrae al cuerpo de los restos, lo que los hace más
pesados y por esto se hunden. Este hecho motiva el desarrollo de sus mandíbulas
inferiores, formando bocas enormes, que pueden recoger restos de organismos,
tanto vegetales como animales, así como estómagos deformables para albergar los
alimentos.
Otra adaptación morfológica es la referente a los sistemas
sensoriales dependientes de la presión como es la línea lateral, donde los
órganos de la misma, o neuromastos se hacen más grandes y presentan canales
amplios.
Los organismos abisales, tienden al gigantismo, que si bien no
se ha podido explicar, la procedencia alóctona del alimento insta a las
criaturas de las profundidades recorrer grandes
distancias, lo que beneficia al tamaño. Otra hipótesis es que debido a las condiciones
extremas en las que viven, crecen muy lentamente, por lo tanto, presentan un
metabolismo lento y longevidad alta.
Como hemos visto, moverse en un medio denso, plantea algunas
dificultades, por lo que los organismos tienden a minimizar los esfuerzos. Una
forma de hacerlo es por medio de reestructurar sus comportamientos, un ejemplo
en este sentido es la conducta reproductiva, donde en algunas en algunas
especies de peces abisales existe dimorfismo sexual
(el macho es mucho más pequeño que la hembra, y vive como parásito pegado a
ella durante toda su vida) reduciendo así el coste de buscar pareja. En
relación al campo de la reproducción, se han adoptado varias tácticas para
reducir la dispersión de los huevos. Los huevos flotantes de los macrúridos de
amplia distribución y de aguas profundas tienen formas escultóricas que frenan
su ascenso, adaptación que no se encuentra en las especies de esta familia que
viven en aguas menos profundas. Los huevos de Hoplostethus atlanticus inicialmente flotan y luego se
hunden, y probablemente terminan su desarrollo en el fondo del mar, con lo que
facilitan su retención en el hábitat adulto.
En lo que se refiere a adaptaciones metabólicas, estas son variadas.
Destacamos significativamente el caso de la fructosa difosfatasa, una enzima
clave para el flujo del carbono durante la gluconeogénesis, que está claramente adaptada para “trabajar mejor”
bajo presión. A concentraciones fisiológicas de enzima y de sustrato, las
velocidades de reacción de las enzimas extraídas del hígado de Salmo gairdneri
(pez que habita en superficie) son muy bajas al ser sometidos a presión,
mientras que las extraídas de hígado de pez cola de rata, Coryphaenoides, que
vive a profundidades de casi 3000 metros, no se afectan por la presión.
4.-Efecto de la hiperbaria sobre el organismo
humano
Como hemos
comentado, dicha variación de presión va a tener una serie de efectos, más o
menos perjudiciales en función del grado de aclimatación del organismo en
estudio. Es por ello, que analizamos los efectos del hiperbarismo desde la
perspectiva de un organismo que
desarrolle su vida a presión atmosférica, recogiendo los efectos de su sometimiento
a presiones elevadas.
Oídos y senos paranasales
Uno de los problemas más comunes a
consecuencia del ascenso de presión, son los barotraumatismos simples del oído medio y en menor medida los
que los afectan al oído interno o a los
senos paranasales.
Si el conducto que conecta el oído
medio y la trompa de Eustaquio no se abre normalmente, la presión en el oído
medio resulta más baja que la del oído externo. En estas circunstancias, el
aumento de presión sobre el tímpano, que separa el oído medio del externo, hace
que éste tienda hacia dentro y, si la presión aumenta hasta cierto punto, se
puede romper, provocar un gran dolor y pérdida de la audición. La entrada de
agua fría en el oído provoca mareos y vómitos.
Las
diferencias de presión en el oído medio pueden afectar al oído interno. Esta presión
desigual genera vértigo (vértigo
alternobárico) cuando se produce el ascenso hacia la superficie.
Las diferencias de presión causan
efectos similares sobre los senos paranasales provocando dolor de cabeza y
cara.
Descompresión.
La enfermedad aguda por descompresión inadecuada (E.A.D.I.) es un
cuadro provocado por fallos técnicos en el ascenso a la superficie después de
un buceo prolongado y profundo.
Las burbujas pueden no aparecer durante
muchos minutos u horas, debido a que los gases pueden permanecer en ocasiones
en un estado de “sobresaturación” durante varias horas antes de formar
burbujas.
La mayoría de los síntomas de la
enfermedad por descompresión son el resultado del bloqueo de los vasos
sanguíneos por burbujas de gas en diferentes tejidos. Inicialmente, sólo los
vasos más pequeños se bloquean por burbujas diminutas, pero a medida que se van
uniendo las burbujas más pequeñas, se van afectando vasos progresivamente
mayores. El resultado es la isquemia tisular y a veces la muerte de los tejidos.
En la mayoría de las personas con
enfermedad por descompresión, los síntomas son dolor en las articulaciones y en
los músculos de las piernas o de los brazos, que afecta al 89% de quienes
padecen enfermedad por descompresión, además de disfunción de la médula
espinal, cerebro y pulmones.
La médula espinal es especialmente
vulnerable, y síntomas aparentemente menores, como debilidad u hormigueo en un
brazo o una pierna, pueden preceder a una parálisis irreversible, a menos que
el proceso se trate de inmediato con oxígeno y recompresión.
El 2% de las personas afectadas por la
enfermedad por descompresión sufren “la asfixia”, causada por cantidades
masivas de microburbujas que taponan los capilares de los pulmones; esto se
caracteriza por disnea grave, seguida con frecuencia de edema pulmonar grave y,
ocasionalmente, por la muerte.
Los efectos tardíos del mal de la
descompresión incluyen destrucción de tejido óseo (osteonecrosis disbárica,
necrosis ósea aséptica), especialmente en el hombro y la cadera, provocando un
persistente dolor y una grave discapacidad.
Se distinguen 3 formas clínicas:
Las tipo I (compromiso cutáneo y/o
osteoarticular);
tipo II (neurológico, respiratorio y/o
cardio-vascular);
tipo III o mixtas (corresponden a
compromisos tipos I y II).
La descompresión provoca daños en los
siguientes órganos:
Dental y óseo (ascenso)
Cerebral (ascenso)
Visceral (ascenso)
·
gastrointestinal
· cavidad
peritoneal
· mediastino
· laringe
· cavidad
craneal
Pulmonar (ascenso)
· enfisema
mediastínico, enfisema subcutáneo y neumotórax
· embolia
gaseosa (de aire)
Otros: enfisema subcutáneo,
barotraumatismo de quiste óseo, afectación de pares
craneales
(V, VII)
Una afección muy común en este sentido
es la embolia gaseosa, la cual consiste en la obstrucción de los vasos
sanguíneos causada por la presencia de burbujas en el flujo sanguíneo,
generalmente producidas por la expansión del aire retenido en los pulmones,
mientras disminuye la presión durante un ascenso. En la aeroembolia, el aire
retenido en los pulmones se expande y los hincha en exceso, produciéndose un
paso de aire hacia el flujo sanguíneo en forma de burbujas. Si éstas obstruyen
los vasos sanguíneos del cerebro, ocasionan daños similares a los que produce
un ataque cerebral grave, como una trombosis o una hemorragia.
Si los pulmones se inflan en exceso
también puede suceder que el aire de su interior llegue a los tejidos que
rodean el corazón (enfisema mediastínico) o incluso bajo la piel (enfisema
subcutáneo). En ocasiones, los pulmones excesivamente cargados revientan, y
liberan aire al espacio que separa a los pulmones de la pared torácica
(neumotórax). En consecuencia, los pulmones se colapsan, produciendo ahogo y
dolor de pecho. Los síntomas que indican que se ha producido una lesión pulmonar
pueden ser la expectoración de sangre o la salida de espuma sanguinolenta por
la boca.
Cambios
del volumen del aire.
Cuando la presión se reduce a la mitad
(como al ascender desde una profundidad de 10 m), el volumen de aire se
duplica. Por ello, si el organismo llena sus pulmones con aire a una
profundidad de 10 m y asciende sin exhalar libremente, el volumen de aire se
duplica, los pulmones se expanden demasiado y puede producirse la muerte, lo
cual se conoce como síndrome de
sobrellenado pulmonar.
Nitrógeno.
Aproximadamente cuatro quintas partes
del aire son nitrógeno. A la presión del nivel del mar, el nitrógeno carece de
efecto conocido sobre la función del organismo, pero a presiones elevadas puede
causar grados variables de narcosis. Cuando el aire es respirado bajo presión,
el nitrógeno (gas inerte) se difunde dentro de los distintos tejidos del
organismo. La captación de nitrógeno por el cuerpo se mantiene mientras la
presión parcial del nitrógeno inspirado sea mayor que la presión parcial del
gas absorbido en los tejidos. La cantidad de nitrógeno absorbido depende de la
presión parcial del nitrógeno inspirado (profundidad) y de la duración de la
exposición. La profundidad a la que aparece una narcosis leve es de unos 36
metros, donde comienza a mostrarse jovial y más despreocupado. A una
profundidad de entre 50 y 60 metros, aparece somnolencia. Entre 60 y 75 metros
el buceador se vuelve casi inútil a consecuencia de la narcosis por nitrógeno.
La narcosis por nitrógeno tiene características similares a las de la
intoxicación alcohólica, y por esta razón se ha denominado el “éxtasis de las
profundidades”.
Intoxicación
aguda por O2.
Cuando la PO2 de la sangre
se eleva muy por encima de 100 mm Hg, la cantidad de oxígeno disuelto en el
agua de la sangre aumenta notablemente. En el intervalo normal de PO2 alveolar,
casi nada del oxígeno total de la sangre es oxígeno disuelto, pero a medida que
la presión aumenta progresivamente hacia los miles de milímetros
mercurio, una gran proporción del oxígeno está disuelto en vez de ligado
a la hemoglobina.
Debido al nivel extremadamente alto de
PO2 tisular que se alcanza cuando se respira oxígeno a una presión
de oxígeno alveolar muy elevada, se producen alteraciones en distintos tejidos.
Este hecho es especialmente acuciado en el caso del encéfalo; una exposición a
4 atmósferas de presión de oxígeno, produciría convulsiones, seguidas de coma.
Otros síntomas que se observan en la intoxicación aguda por el oxígeno son náuseas, sacudidas musculares, mareos, alteraciones
de la visión, irritabilidad y desorientación.
Oxidación de
radicales libres.
Por encima de un nivel crítico de PO2
alveolar (por encima de 2 atmósferas de PO2), fracasa el sistema
amortiguador de hemoglobina/ oxígeno, y la PO2 tisular puede elevarse
a cientos o miles de milímetros de mercurio. Entonces la cantidad de radicales
libres oxidantes sobrepasa la capacidad de los sistemas enzimáticos encargados
de eliminarlos y ejercen efectos gravemente destructivos e incluso letales
sobre las células. Uno de los efectos principales es oxidar los ácidos grasos
poliinsaturados, que son un componente esencial de muchas de las estructuras
membranosas de las células. Los tejidos nerviosos son especialmente sensibles
por su gran contenido en lípidos. Por tanto, buena parte de los efectos letales
agudos de la toxicidad del oxígeno están relacionados con la disfunción
cerebral, no obstante, antes de que se llegue a estos extremos habrá aparecido
daños en otras estructuras motivadas por otros efectos de la presión.
CO2.
Los valores anormalmente altos de
anhídrido carbónico (intoxicación por anhídrido carbónico) pueden causar
pérdidas transitorias de la visión y de la conciencia. Algunas personas
presentan una acumulación de anhídrido carbónico porque no aumentan su
frecuencia respiratoria adecuadamente durante el esfuerzo físico. Las altas
concentraciones de anhídrido carbónico aumentan la posibilidad de que se
produzcan convulsiones secundarias a la toxicidad del oxígeno e incrementan la
gravedad de la narcosis del nitrógeno.
A nivel celular.
A nivel
celular, las altas presiones provocan inhibición de la síntesis de RNA, DNA y
proteínas, mal funcionamiento
en el transporte de membrana, disminución de la tasa de actividad de varias
enzimas y la posible distorsión de la configuración
terciaria de las proteínas.
5.- Medicina hiperbárica
La
MEDICINA HIPERBÁRICA es la rama de la ciencia que estudia los cambios
fisiológicos y fisiopatológicos de los seres vivos sometidos a presiones
superiores que la atmosférica.
•
Qué es la Oxigenación Hiperbárica ?
La
terapia de Oxigenación Hiperbárica es un método de tratamiento médico en donde
se somete al cuerpo entero a una presión parcial de oxígeno mayor que la
atmosférica y el paciente respira oxígeno al 100% en el interior de un
recipiente denominado: Cámara Hiperbárica.
•
Beneficios :
-
Reduce la hipoxia tisular
-
Mejora el sistema inmune
-
Acelera el proceso de cicatrización
-
Apoya la regeneración ósea
-
Efectos adversos ocasionados por la
Oxigenoterapia Hiperbárica.
-
miopía reversible
-
cataratas
-
rotura del oído medio
-
sinusitis
-
lesiones en dientes y tejido pulmonar
-
crisis epilépticas
No hay comentarios:
Publicar un comentario